
Marina Zieger University of Michigan, Department of Ophthalmology and Visual Sciences, Ann Arbor, USA
The cephalic image-forming eyes in Lymnaea stagnalis are important distant sensory organs that continuously inform the CNS about the intensity and direction of light stimuli (Zieger and Meyer-Rochow, 2008).
However, how photoreceptors in the eye of L. stagnalis are organized, how they respond to light stimulation, how they send afferent information to the CNS, and additionally how they receive efferent information from the CNS are not fully understood.
Recent intracellular recordings using electrodes filled with a tracer dye (Lucifer yellow) revealed that there are at least two types of light-sensitive cells, type A and type T, which have distinct light sensitivity, and morphological features in the L. stagnalis eye (Sakakibara et al., 2005). The authors demonstrated that the type A cells do not send, whereas the type T cells do send their axons to the optic nerve and to the CNS; type T cells are more light sensitive than the type A cells in both absolute and dynamic sensitivity; type A cells send axons to the type T cells within the retina.
My personal anatomical and ultrastructural studies (Bobkova, 1998; Bobkova et al., 2004) suggested that there are two morphological classes of photoreceptors, termed type I (the cells with finger-like apex, bearing long light-sensitive microvilli) and type II (cells with convex apex and short tuff of microvilli) in the L. stagnalis eye.
My attempt to match the remarkable results obtained by Sakakibara et al. (2005) and my own structural observations led me to initiate the additional morphological study.
Proposed research
The research aims at combining our earlier achievements in the understanding of the cellular compositions of the retina and the optic nerve in L. stagnalis, with new findings on interactions between the elements of the eye-CNS network.
Questions:
1. How many types of retinal cells are there in the eye and which of them contribute to the optic nerve?
2. What are the functions of gap junctions, and synapse-like various morphology contacts between retinal cells and their axons?
3. What are the composition of the plexiform layer, both neuropil and the distal portions of the optic nerve, as well as the composition of the eye capsule?
4. What is a topology of the efferent innervation?
Research to date
1. Our studies demonstrate that most of the photoreceptors appear to be neurobiotin-filled. A beaded appearance of the marker along nerve fibers suggests multiple synaptic varicosities. No terminal arborizations of photoreceptors were revealed within the plexiform layer of the retina. Therefore, the retrograde transport of neurobiotin shows that both type I and type II photoreceptor cells send their axon to the optic nerve.
Since microvilli are considered to be the photoreceptive region, the incident light must be absorbed much more effectively by type I cells than by type II cells, suggesting that the former must be more sensitive to light than the latter. It is reasonable to assume that type I and type II cells in our research correspond to type T and type A cells, respectively, in the Sakakibara et al. study (2005).
Tracing studies using cobalt chloride and horseradish peroxidase back-fill methods identified a population of retinal neurons that send their axons to the optic nerve. The retinal neurons have no specialized light-receiving membrane, have large nucleus and characterized by presence of large dense-core vesicles and do not form any synapse-like contacts within the retina. They are located deep in the somatic and plexiform layers of the retina and generally are fewer in number (10 to 15) compare to the specialized photoreceptors that are numerous and equally distributed in the retina. It is possible that the neurons belong to the sensory-neurosecretory type (Zieger et al., 2009; Tessmar-Raible et al., 2007) or play a role as circadian pacemakers (Olson and Jacklet, 1985).
2. Ultrastructural observations reveal the multiple contact sites between the axons within the plexiform layer of the retina. Most of the axon-axonal contacts are symmetrical, with clear or dense vesicles and premembranous densification on both sides of the cleft. Most likely at these junctions, transmission may occur in both directions across the synapse.
Structural studies confirm the existence of gap junctions suggested by Sakakibara et al. (2005) between photoreceptors. It is possible that there is an electrical coupling between type I and type II photoreceptors, however the functional role of it remains to be understood.
3. Each eye projects to the ipsilateral cerebral hemiganglion via a long (up-to 1 cm) optic nerve, which contains about 2800 axons of 0.5 μm and about 200 axons of 1.5 μm in diameter. The thin axons belong to the photoreceptor cells and less than 10% of the thick axons belong to the retinal neurons. Throughout most of the examined distal portion of the optic nerve, there was no well-defined synaptic interaction between the axons. The lateral terminal projections of the neurobiotin-filled fibers in the optic nerve form a glomerus-like structure in the neighbouring tentacular nerve.
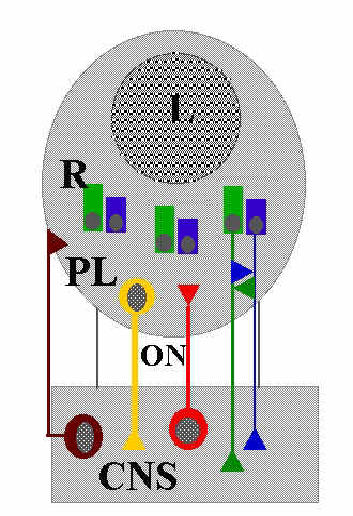 |
| Figure 1: Summary diagram of the eye network of Lymnaea stagnalis. Photoreceptors of type I (green), photoreceptors of type II (blue), retinal neurons (yellow), serotonergic efferent innervation of the retina (R) (red) via optic nerve (ON) and efferent innertation of the eye capsule (brown), lens (L), as well as the central nervous system (CNS) are indicated. Triangles mark sites of synaptic interactions. |
4. The immunohistochemical labelling of the efferent sensory pathways within the optic nerve and eye shows serotonergic efferent innervation of the eye capsule. Many efferent serotonergic fibers terminate in the plexiform layer suggesting projections of the central neurons into the retina through the optic nerve.
Conclusions and Significance
The important conclusions from these studies are that the retina is composed of two types of neurons: photoreceptors and retinal neurons; photoreceptors are united into an morphological network and may be coupled electrically; the central neurons send their projections into the eye via the optic nerve and the eye capsule is innervated through both the optic nerve and separate nerve branches (Fig. 1).
Thus, Lymnaea stagnalis can now be used as a model system to examine the influence of the central projections of the identified retinal neurons on the animal behaviour.
The funding from Malacological Society of London allowed me to continue my research on molluscs, and I am extremely grateful for the financial assistance.
Literature Cited
Bobkova, M.V. 1998. Structural and functional organization of the peripheral part of the visual system of the common pond snail Lymnaea stagnalis . Journal of Evolutionary Biochemistry and Physiology 34, 531-546.
Bobkova , M.V., Gál, J., Zhukov, V.V., Shepeleva, I. P., and Meyer-Rochow, V. B. 2004. Variations in the retinal designs of pulmonate snails (Mollusca, Gastropoda): squaring phylogenetic background and ecophysiological needs (I). Invertebrate Biology 123 (2), 101-115.
Olson, L. M., and Jacklet, J. W. 1985. The circadian pacemaker in the Aplysia eye sends axons throughout the central nervous system. Journal of Neuroscience 5, 3214-3227.
Sakakibara, M., T. Aritaka, A. Iizuka, H. Suzuki, and T. Horikoshi. 2005. Electrophysiological responces to light of neurons in the eye and statocyst of Lymnaea stagnalis. J Neurophysiol. 93, 493-507.
Tessmar-Raible, K., Raible, F., Christodoulou, F., Guy, K., Rembold, M., Hausen, H., and Arendt, D. 2007. Conserved sensory-neurosecretory cell types in annelid and fish forebrain: Insights into hypothalamus evolution. Cell 129, 1389-1400.
Zieger, M., and Meyer-Rochow, V. B. 2008. Understanding the cephalic eyes of pulmonate gastropods: A review. American Malacological Bulletin 26, 47-66.
Zieger M. V. , Vakoliuk I. A., Tuchina O. P., Zhukov V. V., and Meyer-Rochow V. B. 2009. Eyes and vision in Arion rufus and Deroceras agreste (Mollusca; Gastropoda; Pulmonata): What role does photoreception play in the orientation of these terrestrial slugs? Acta Zoologica (Stockholm) 90, 189-204.

Indrella amputta. © The Natural History Museum, London.

Mariaella dussumieri. © The Natural History Museum, London.
. © The Natural History Museum, London.")
Beddomea trifasciatus (lowland form). © The Natural History Museum, London.