Morphological and molecular study of the geminate species of
opisthobranch mollusks from the Central American isthmus.
Yolanda E. Camacho-Garcia,
Museo de Zoologia, Escuela de Biologia, Universidad de Costa Rica
"Geminate species” are species that occur on different coasts
of the Central American isthmus but are very similar morphologically.
They are used to infer rates of molecular evolution (Fig.
1) (Marko, 2002). Based on the assumption that species were isolated
geographically by the latest stages of the seaway closure 3.1-3.5 million
years ago (Coates and Obando, 1996), the amount of sequence divergence
between germinate species can be used to estimate the rates at which
genes diverge (Knowlton et al., 1993). Valdes (2001) suggested that
before the closure of the Central American isthmus, some opisthobranch
species were potentially distributed throughout the Caribbean and eastern
Pacific, and that the rising of the Panama land bridge caused fragmentation
of the range and subsequent speciation. This vicariance hypothesis,
which seems to explain the apparent close relationship between the Caribbean
and the eastern Pacific faunas, has been tested on a variety of marine
invertebrates (Vermeij, 1978; Lessios and Weinberg, 1994; Lessios et
al., 1999), but only on a small clade of opisthobranchs (Gosliner and
Johnson, 1999). It has never been dated using molecular techniques.
Proposed research
In my grant application I proposed to test the vicariance hypothesis
using a combination of molecular and morphological characters to identify
geminate species in opisthobranch molluscs from the Atlantic and Pacific
coasts of Costa Rica and Panama, and compare the results with these
on other marine invertebrates. There were three objectives in my study:
Objective 1: To document the biodiversity of opisthobranch molluscs at both coasts of Costa Rica and compare the faunal composition.
Objective 2: To identify and characterize geminate species pairs and test the molecular clock using a combination of morphological and molecular characters (i.e. cytochrome oxidase I and H3 genes).
Objective 3: To determine whether there is a genetic
divergence among different populations/species of circumtropical opisthobranchs
at both sides of the Panama isthmus.
Research to date
Objective 1: A large-scale and ongoing inventory was
conducted at the Atlantic and Pacific coasts of Costa Rica (Manzanillo,
Pto. Viejo, and Cahuita and multiple localities in Guanacaste and the
Osa Peninsula, respectively), and Panama [Bocas del Toro (trip supported
by this grant, awarded in 2006) and Isla Coiba, respectively]. Traditional
sampling methods (i.e. tidepooling, snorkeling and scuba diving) were
used. A number of additional samples, collected during separate expeditions
in the Indo-Pacific and South Pacific, have also been taken into account.
New records from both coasts are reported for the first time (Hermosillo
and Camacho-Garcia, 2006). Several new species are also described (a
new species of Janolus: Camacho-Garcia and Gosliner, 2006;
new species of the genera Jorumna and Paradoris (Camacho-Garcia
and Gosliner, in press) and of the genera Atagema and Taringa.
The study documents 250 species of opisthobranch molluscs, of which
127 are found only in the Pacific and 111 only in the Caribbean coasts
(Camacho-Garcia, in press). According to this inventory the Pacific
coast has a higher number of nudibranch dorids and aeolids than the
Caribbean; however, at both coasts the sponge eaters dominate.
The closure of the Panama isthmus (about 3.1 million years ago) has
prevented any contact among opisthobranch species from the two coasts
of Costa Rica (Coates, 1997). However, there are recorded 12 species
in common in the Pacific and the Caribbean coasts. Examples include
Flabellina marcusorum, Oleurobrancus areolatus, Lobiger
souverbii. Most of these species are found worldwide in tropical
areas, e.g. Stylocheilus striatus and Aplysia parvula
(Fig. 2).
Objectives 2 and 3: A phylogenetic analysis of all
the taxa studied in objective 1 is planned. To clarify the relationships
among geminate opisthobranch species I have used an approximately 700bp
fragment of the cytochrome oxidase I gene (COI; primers by Folmer et
al., 1994), and I intend to incorporate new data based on an amplification
of a segment of 274bp of the H3 gene. This work is currently being performed
at the Osher Laboratory, California Academy of Sciences (CAS), and is
supported by a PEET grant (Partnerships for Enhancing Expertise in Taxonomy
from the National Science Foundation, USA) to T. Gosliner and A. Valdes.
So far, more than 200 sequences have been proofread. The completion
of this work, including analysis of data and evaluation and presentation
of biogeographical implications of the findings, is due to be completed
after a second, forthcoming visit of mine to CAS.
Literature cited
| Camacho-Garcia, Y., in press. Benthic Opisthobranchs from Costa Rica (Mollusca: Gastropoda). In | |
| (Ed.) I Wehrtmann and J. Cortes, Marine Biodiversity of Central America: The case of Costa Rica Kluwer Academic Press | |
| Camacho-Garcia, Y. and T. Gosliner, in press. The genus Paradoris Bergh, 1884 (Nudibranchia: | |
| Discodorididae) from the Tropical Americas. The Veliger 49(3). | |
| Camacho-Garcia, Y. and T. Gosliner, in press. Systematic revision and morphological phylogenetic | |
| analysis of the genus Jorumna Bergh, 1876 (Nudibranchia: Discodorididae). Journal of Molluscan Studies. | |
| Camacho-Garcia, Y. and T. Gosliner, 2006. New species of zephyrinid genus Janolus Bergh, 1884 | |
| (Opisthobranchia:Nudibranchia) from North America and Costa Rica. Revista de Biologia Tropical 54(4): 1295-1305. | |
| Coates, A.G. and J.A. Obando, 1996. The geologic evolution of the Central American Isthums. In | |
| (Ed.) Jackson, JBC, AF Budd and AG Coates, Evolution and Environment in Tropical America The University of Chicago Press, Chicago, | |
| Folmer, O., M. Black, W. Hoeh, R. Lutz and R. Vrijenhoek, 1994. DNA primers for amplification of | |
| mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3: 294-299. | |
| Gosliner, T.M. and R. Johnson, 1999. Phylogeny of Hepselodoris (Nudibranchia: Chromodorididae) | |
| with a review of the monophyletic clade of Indo-Pacific species, including descriptions of twelve new species. Zoological Journal of the Linnean Society, London125: 1-114. | |
| Hermosillo, A. And Y. Camacho-Garcia, 2006. A note on the opisthobrancs of Parque Nacional de | |
| Coiba, Panama (tropical Eastern Pacific). The Festivus 38(9): 95-98. | |
| Knowlton, N., L.A. Weigt, L.A. Solorzano, D.K. Mills and E. Bermingham, 1993. Divergence in | |
| proteins, mitochondrial DNA, and reproductive compatibility across the Isthmus of Panama. Science 260: 1629-1633 | |
| Lessios, H.A. and J. Weinberg, 1994. Genetic and morphological divergence among morphotypes of | |
| the isopod Excirolana on the two sides of the Isthmus of Panama. Evolution 48: 530-548 | |
| Lessios, H.A., B.D. Kessing, D.R. Robertson and G. Paulay, 1999. Phylogeography of the pantropical | |
| sea urchin Eucidaris in relation to land barriers and ocean currents. Evolution 53: 806-817 | |
| Marko, P. 2002. Calibration of molecular clocks and divergence times of geminate species pairs | |
| separated by the Isthmus of Panama. Molecular Biology and Evolution 19: 2005-2021 | |
| Vermeij, G. 1978. Biogeography and adaptation Harvard University Press, Cambridge | |
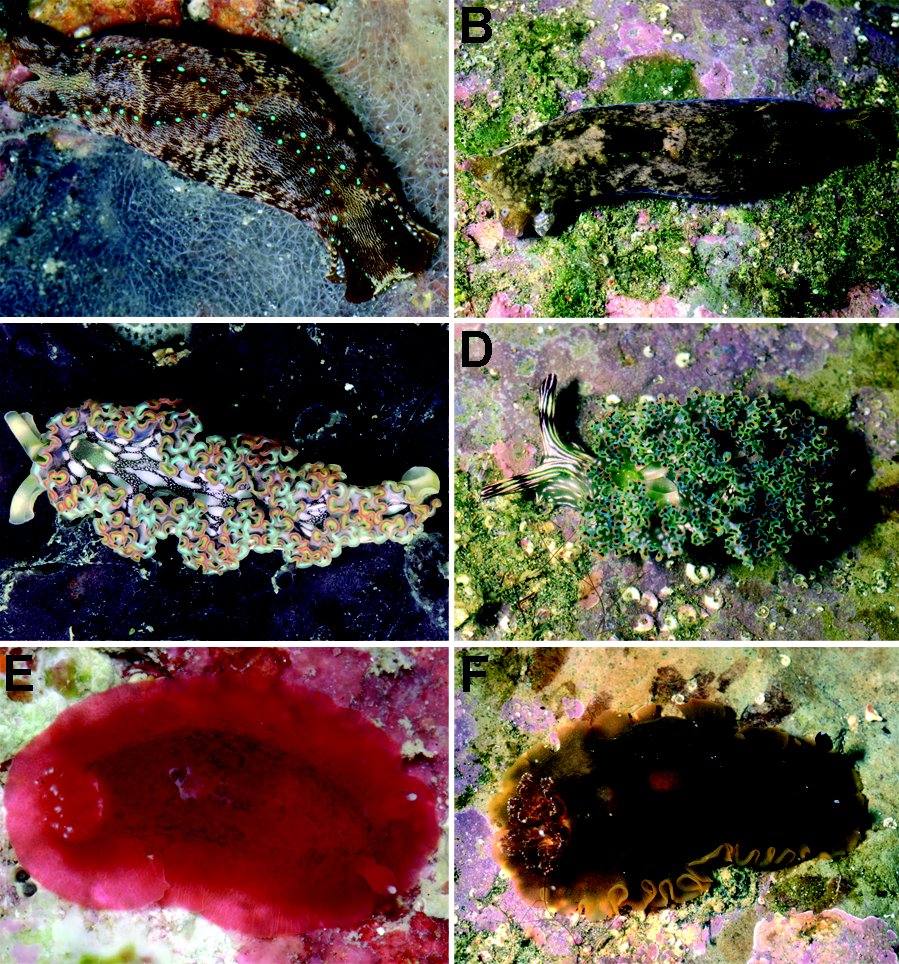
Figure 1: Potential geminate species of opisthobranch molluscs. A. *Navanax aenigmaticus, (Caribbean) B. **Navanax aenigmaticus (Pacific). C. *Elysia crispata (Caribbean). D. **Elysia diomedea (Pacific). E. *Dendrodoris krebsii (Caribbean). F. **Dendrodoris fumata (Pacific), (*): photo by A. Valdés, (**): photo by T. Gosliner.
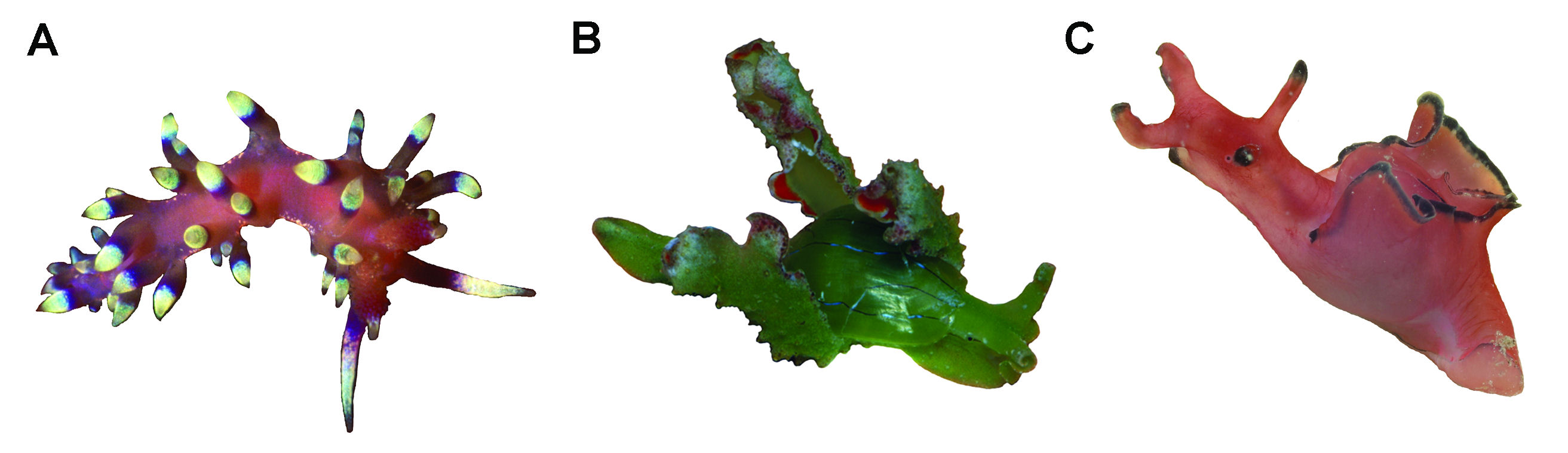
Figure 2: Examples of species present along the Central American isthmus. A. Flabellina marcusorum, photo by T. Gosliner. B. Aplysia parvula, photo by Y. Camacho. C. Lobiger souverbii, photo by A. Valdés.

Ammonite
")
Glaucus marginatus Bergh, 1860. © D. Riek (http://www.roboastra.com/)
. © The Natural History Museum, London.")
Beddomea trifasciatus (lowland form). © The Natural History Museum, London.