
Roland Schultheiß, Justus Liebig University,
Giessen, Germany
Ancient lakes, i.e. lakes with an estimated age of
over 100,000 years, are remarkable hotspots of biodiversity. Research
on the evolution and phylogeny of the fauna of these lakes, however,
has mainly concentrated on conspicuous taxa like the colourful cichlid
fish in the African lakes Victoria and Malawi or the so called thalassoid
(i.e., shells with strong ribs and spines) gastropods of Lake Tanganyika
(African Rift Valley) or Lake Sulawesi (SE Asia). More inconspicuous
taxa, on the other hand, remained out of focus although the contribution
of these groups to the biodiversity of ancient lakes is believed to
be considerable. A case in point is the pea clam genus Pisidium
C. Pfeiffer, 1821 (Bivalvia, Sphaeriidae): due to the scarcity of morphological
characters, determination on species level is difficult and requires
a considerable degree of taxonomic experience. Endemic species of this
'notoriously difficult genus of freshwater bivalves' (Dance & Kuiper,
2002) are reported from ancient lakes only.
In a world-wide context, the European ancient lakes Ohrid and Prespa
harbour an exceptional pea clam fauna: one endemic species is reported
from each lake. Altogether, both lakes are inhabited by nine Pisidium
species (Korniushin, 2004) (Fig. 1). Hypotheses
on the evolution of the two endemic species as well as the underlying
phylogenetic relationships were so far based on morphological and anatomical
data only (Kuiper, 1960; Kuiper, 1987; Korniushin et al., 2000). The
consequential need of molecular based analyses of the pea clam fauna
of the lakes Ohrid and Prespa was stressed already by Korniushin (2004).
The project 'Systematics and character evolution of Pisidium
(Bivalvia) in the ancient lakes Ohrid and Prespa' aims to close this
gap. Using DNA sequence data as well as a morphometrical approach based
on standard shell parameters and shell shape analyses:
Objective 1: We attempted to reveal the underlying relationships
of the pea clam fauna in the two ancient lakes. By including Pisidium
species from all over Europe, North America and Asia in our dataset,
we were able to establish a broad phylogenetic framework.
Objective 2: We tested for phylogenetic signals within the
morphometrical dataset and compared it to the molecular ones.
Objective 3: We embedded our findings in a European biogeographical
context and proposed scenarios on the evolution of the endemic pea clam
species in the lakes Ohrid and Prespa.
Results & Discussion
Objective 1: Based on the mitochondrial 16S (large ribosomal
subunit) fragment, we conducted a phylogenetic analysis using Bayesian
inference as implemented in the program MrBayes 3.0b3 (Huelsenbeck and
Ronquist, 2001). Our dataset comprised all reported pea clam species
of the lakes Ohrid and Prespa (with exception of P. moitessierianum
of which we have found only a single valve in three years). Furthermore,
we included nearly all European Pisidium species in our phylogeny
(with exception of P. conventus, P. dilatatum, P. hinzi, P. pulchellum
& P. sogdianum) as well as several North American and Asian
species. Additionally, we performed a parsimony-based network analysis.
Our analyses revealed the existence of two cryptic lineages within the
pea clam fauna of the two ancient lakes. Members of the two lineages
were previously determined as P. casertanum. In accordance
with the findings of Lee and Ó'Foighil (2003), we found a paraphyletic
species complex of P. casertanum. A new cryptic lineage, likewise
originally determined as P. casertanum, appears to be only
very distantly related to this complex. Furthermore, we were able to
test the hypothesis of a sister relationship of the two endemic species
P. edlaueri (Lake Ohrid) and P. maasseni (Lake Prespa).
This relationship was confirmed on the basis of molecular data. It has
to be pointed out, however, that the relationship appears to be so close
that the species status of both endemics might be in question.
Objective 2: Using elliptic Fourier analyses we conducted principle component analyses (PCA) on the basis of shell outline data. Neither a PCA nor a discriminant analysis could separate the analysed pea clams on species level. We then conducted a mantel test in order to assess whether the evaluated morphometrical distances within Pisidium correspond to genetic distances. The result was a weak but highly significant positive correlation between both datasets (r = 0.32; p = 0.001). This confirms the findings of Guralnick (2005), who suggested that lineage specific shell patterns are not completely masked by eco-phenotypic variance.
Objective 3: Within a European context, pea clam fauna in the lakes Ohrid and Prespa is remarkable (e.g. Kuiper, 1960) in respect of species number and composition. In the present study we addressed the latter phenomenon by testing the hypothesis on the origin of the endemics P. edlaueri and P. maasseni. Kuiper (1960) regarded them as variants of P. nitidum, a species that does not occur within the two lakes. This was also confirmed by the present study. Genetically, the three taxa were, however, so closely related that either all three taxa belong to one species or the resolution of the 16S fragment was not sufficient for resolving the relationship. On the basis of genetic diversity and network structure, we propose a "stepping stone" scenario of colonisation of the two lakes by the most recent common ancestor (MRCA) of the recent endemic species: therewith, the MRCA first invaded Lake Ohrid and used this lake as stepping stone for the subsequent colonisation of Lake Prespa.
Conclusion
Our study stresses the importance of taxonomic expertise, particular
in notorious taxa like Pisidium. Since modern morphometrical
analyses fail to provide straight forward characters for species determination,
considerable taxonomic experience is required to work with such difficult
taxa. This expertise - combined with ecological familiarity with the
taxon in question - forms the framework for further research on the
subject. Only on that basis, the application of modern approaches like
DNA sequencing might be reasonable and efficient.
Literature cited
| Dance, S.P. and J.G.J. Kuiper, 2002. B.B. Woodward and the 'Pisidium affair'. Journal of Conchology 37: | |
| 635-650. | |
| Guralnick, R.P., 2005. Combined molecular and morphological approaches to documenting regional | |
| biodiversity and ecological patterns in problematic taxa: a case study in the bivalve group Cyclocalyx (Sphaeriidae, Bivalvia) from western North America. Zoologica Scripta 34: 469-482. | |
| Huelsenbeck, J.P. and F. Ronquist, 2001. MrBayes: Bayesian inference of phylogeny. Bioinformatics | |
| 17: 754-755. | |
| Korniushin, A.V., 2004. The bivalve mollusca fauna of ancient lakes in the context of the historical | |
| biogeography of the Balkan region. In (Ed) H.I. Griffiths, B. Krystufek and J. M. Reed Balkan Biodiversity - Pattern and Process in the European Hotspot. Kluwer Academic Publishers, Dordrecht/Bosten/London, | |
| Korniushin, A.V., Z. Krstanovski and G. Kostoski, 2000. Anatomical evidence of close affinity between | |
| endemic species of Pisidium (Bivalvia, Sphaeriidae) from some ancient lakes, and the widely distributed taxa. Journal of Zoological Systematics and Evolutionary Research 38: 81-86. | |
| Kuiper, J.G.J., 1960. Die Pisidien des Ohridsees, Mazedonien, nebst Bemerkungen über die | |
| Verbreitung der Pisidien in der Balkanhalbinsel und den Donauländern. Beaufortia 7: 219-231. | |
| Kuiper, J.G.J., 1987. Pisidium maasseni n. sp., a new species from Lake Prespa, Jugoslavia (Bivalvia, | |
| Sphaeriidae). Basteria 51: 163-165. | |
| Lee, T. and D.Ó. Foighil, 2003. Phylogenetic structure of the Sphaeriinae, a global clade of freshwater | |
| bivalve molluscs, inferred from nuclear (ITS-1) and mitochondrial (16S) ribosomal gene sequences. Zoological Journal of The Linnean Society 137: 245-260. | |
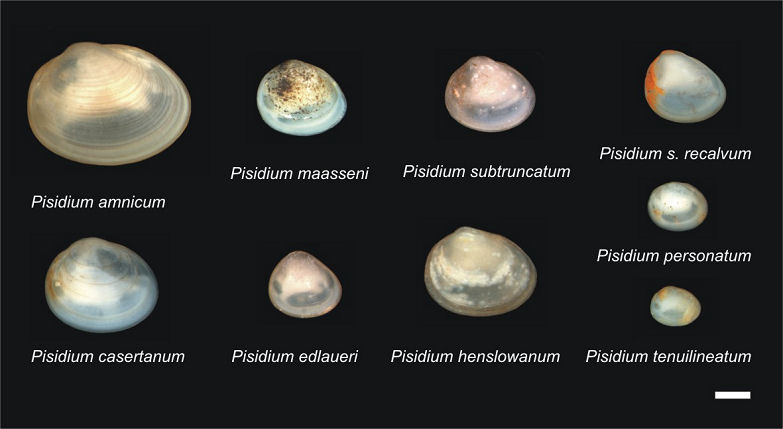
Pisidium species of the ancient lakes Ohrid and Prespa (with
exception of P. moitessierianum; see discussion for details).
P. edlaueri is endemic to Lake Ohrid while the closely related
P. maasseni inhabits Lake Prespa. Additionally, from Lake Ohrid
an endemic subspecies, P. subtruncatum recalvum, is reported.

Indrella amputta. © The Natural History Museum, London.

Mariaella dussumieri. © The Natural History Museum, London.
. © The Natural History Museum, London.")
Beddomea trifasciatus (lowland form). © The Natural History Museum, London.