The meeting of The Malacological Society of London in the Department of Earth Sciences, Open University, Milton Keynes Wednesday 5th June 2002, organised by Professor Peter Skelton
Evolution of polar molluscan faunas J.
Alistair Crame. British Antarctic Survey, High Cross, Madingley
Road, Cambridge CB3 0ET. Various molluscan taxa have been instrumental in formulating our concept of large- scale biogeographical patterns in the marine realm. On perhaps the largest scale of all, it is apparent that high-latitude and polar molluscan faunas can be traced back from the present day to at least the Late Palaeozoic era. Their presence points to the existence of some form of global-scale faunal differentiation: tropical biotas have in some way been consistently separated from temperate and polar ones. For many observers an explanation of such phenomena lies in a fundamental difference in evolutionary rates. Low-latitude and tropical faunas are thought to have higher rates of radiation (speciation), and high-latitude and polar ones higher rates of extinction. Evidence will be presented to suggest that, whilst the former of these statements can be upheld, the latter cannot. We still have much to learn about rates of molluscan evolution on a global scale.
Mid-Aptian demise of Tethyan carbonate platforms: a role for climate? Peter Skelton. Department of Earth Sciences, The Open University, Milton Keynes MK7 6AA The extensive carbonate platforms of the Cretaceous greenhouse world showed episodic development, punctuated by crises of regional to global extent. These crises also involved mass extinctions among the benthic platform biota, especially rudists. The most prominent examples occurred in the mid-Aptian and the terminal Cenomanian, the former of which is investigated here. Various scenarios involving linkage between climate, oceanic anoxia and the global carbon cycle have been postulated, though detailed stratigraphical resolution of the pattern of events is still needed so that likely cause and effect can be disentangled. Current data for the timing of Tethyan platform terminations and associated rudist extinctions show a trend of regional restriction to lower palaeolatitudes (with perhaps a West to East component) extending from before, to the end of Bowerbanki Zone times: the latest Bedoulian developments occurred on the central and southern platforms of eastern Tethys (e.g. the Shu'aiba Formation of Arabia). Early Gargasian deposits suggest wetter (?cooler) conditions in many former platform areas. Recovery of the rudist fauna was fairly slow, with widespread platform development only taking off again in the latest Aptian and Albian. The duration and regional pattern of these changes suggest relatively long-term climatic influences. Given the huge quantities of carbonate carbon that were sunk in the platform reservoirs, one possible hypothesis is that the growth of the platforms, coupled with an irregular volcanic supply of atmospheric carbon dioxide, fed back to the climate change that led to their own demise. Polar molluscan vent faunas: examples from the Jurassic of the Antarctic and the Cretaceous of Greenland Simon R.A. Kelly. CASP, University of Cambridge, West Building, 181A Huntingdon Road, Cambridge CB3 0DH. Email: simo...@casp.cam.ac.uk Two molluscan vent faunas are discussed: one from the Atoll Nunataks Formation, Fossil Bluff Group (Tithonian, Late Jurassic) of Alexander Island, Antarctic Peninsula; the other from the Kuhnpasset Beds (Barremian, Early Cretaceous) of Wollaston Forland, Northeast Greenland. Although both faunas are associated with anomalous high latitude limestones, in otherwise deep water mudstones, the molluscan faunas of these vents are very different and there are contrasts between the limestones. The Antarctic fauna is dominated by a small, as yet unidentified, gastropod, with occasional large bivalves (?Cryptolucina); also present are crinoids, foraminiferans and ostracods, and rare ammonites (Lytoceras) and belemnites (Belemnopsis). Associated calcareous laminated crusts bordered near- or at-surface cavities, and show radial-fibrous and botrioidal carbonate cements. Very negative carbon isotope values demonstrate 13C values of -33.5A to -44.6A PDB which indicate methane oxidation for the carbonate. The limestone body is a lens about 200 m wide and up to 3 m in thickness. It is the only limestone in a c10 km thick forearc basin succession. Brecciated limestone blocks suggest methane ponding at subsurface followed by explosive release resulting in a pockmark structure (Kelly et al. 1995). The Greenland vents are dominated by large bivalves. These include an abundant lucinid, Cryptolucina kuhnpassetensis Kelly, a common large modiomorphid, Caspiconcha whithami Kelly which reaches over 300 mm in length and has a shell up to 28 mm in thickness, and less commonly, Solemya. Also the wood-boring bivalve Turnus is abundant in common driftwood. Gastropods are rare, but the associated cephalopod fauna includes ammonites (Lytoceras and heteromorphs), belemnites (Oxyteuthis), nautiloids (Cymatoceras) and a remarkable large orthoconic phragmocone. They occur in carbonate mounds over a stratigraphic interval of 150 m and crop out over a 1.5 x 0.5 km area. The limestone takes the form of over 30 mounds up to 2 m high and 3-4 m wide. They show carbonate-cemented tube systems, associated laminated calcite crusts and void fills. The geochemistry of the carbonates is still undergoing analysis, so it cannot yet be confirmed that methane is the origin of the vents. However, whatever the source, it was ephemeral and fluid movement routes through the sediment changed frequently (Kelly et al. 2000). The contrasts between the two vents may be ascribed to a number of factors including: Rates of nutrient flow. This controls the form of the limestone body: fast flow gives raised calcareous chimneys; moderate flow gives low topography mounds, with associated shallow-depth localised cementing; slow flow may result in subsurface ponding followed by explosive release. Recruitment of new fauna. Sites can only be colonised if there is an up-current site already established. Advantage of lecithotropic larvae. Type of feeding. Regardless of the actual nutrient, feeding is highly specialised. The gastropod-dominated fauna was likely feeding as an algal grazer on the probable bacterial origin carbonate crusts. Whereas the bivalves such as Cryptolucina and Caspiconcha were probably forms with modified gills, which bore bacterial 'farms', from which these 'gutless' bivalves obtained their nutrition. The presence of nektonic feeders is believed to be opportunistic discovery by cephalopods.
Hydrothermal vent and cold seep molluscs: view from the fossil record Crispin T. S. Little and Kathleen A. Campbell School
of Earth Sciences, University of Leeds, Leeds LS2 9JT. Email:
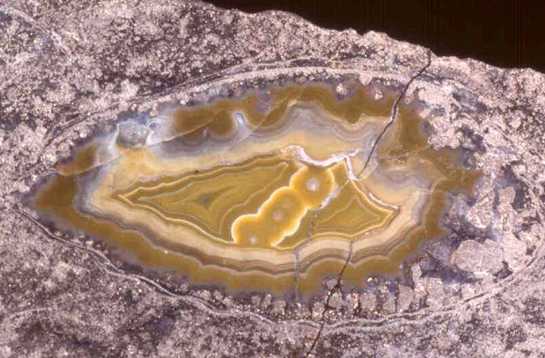
c.li...@earth.leeds.ac.uk Email: ka.c...@auckland.ac.nz In the presentation we will review the occurrence of molluscs in ancient hydrothermal vent and cold seep communities, discuss the role of chemosymbiosis in these communities, and map out future work in the field. The entry of molluscs into vent and seep environments occurred early in the Palaeozoic, as a limited diversity of molluscs (monoplacophorans, bivalves and gastropods) is present in the oldest known vent (Silurian, ~430 Ma) and seep (Devonian, ~380 Ma) fossil assemblages. A few Palaeozoic vent and seep assemblages are dominated by bivalves, but others contain no molluscs, or are outnumbered numerically by brachiopods. Due to poor preservation many Palaeozoic vent and seep molluscs are difficult to identify. Others belong to extinct groups, common in contemporaneous non-vent and non-seep fossil assemblages. These factors do not allow chemosymbiotic lifestyles to be established with any certainty for these ancient taxa. The record is better in the Mesozoic and Cenozoic, especially for seeps. Towards the later Mesozoic, brachiopods become increasingly rare in vent and seep communities, and bivalves and gastropods become the dominant shelly taxa, both in abundance and diversity. The Mesozoic marks the first appearance in vents and seeps of a group of chemosymbiotic bivalves (mytilids, lucinids, and solemyids) which are important constituents of modern chemosynthetic communities, but also have Palaeozoic, non-vent, non-seep origins. The first provannid gastropods and vesicomyid bivalves appear in late Jurassic and early Cretaceous seeps, respectively. These two families are today restricted to chemosynthetic communities and have many chemosymbiotic species. Many modern vent and seep molluscs, particularly the smaller gastropods, have yet to be identified in the fossil record, although this pattern may change as taxonomic work progresses on recent gastropod finds at Mesozoic and Cenozoic seep and vent sites. In summary, Phanerozoic vent and seep deposits contain a rich and growing record of fossil molluscs, which serves as the litmus test for comparison of phylogenetic hypotheses generated by molecular data from living chemosymbiotic molluscs. Figure Cross section through Anarhynchia, a rhynchonellid brachiopod filled with agate from the early Jurassic aged Figueroa vent site in Santa Barbara County.
Whatever Happened to the Shelled Cephalopods? Neale Monks Depít Palaeontology, Natural History Museum, London. Email: N.Mo...@appleonline.net As victims of the end-Cretaceous mass extinctions, the demise of the ammonites and the belemnites is well known. What is less widely appreciated is that as modes of life, the ammonite niche and the belemnite niche appear to have persisted well into the Tertiary. Post-Cretaceous nautiloids are abundant and widespread, and many are distinctly ammonite-like, with complex, rather than simple, sutures and a variety of coiling modes and even ornamentation. Similarly, coleoids with robust internal shells with gas-filled chambers converge morphologically with belemnites. These ammonite-like nautiloids and belemnite-like coleoids lived alongside the more conservative nautiloids on the one hand and the progressive coleoids with reduced or absent shells on the other throughout much of the Tertiary. By the time of the Miocene, however, cephalopod diversity was again reduced, and in modern seas there are no ammonite-like nautiloids at all, and only one species of highly modified coleoid, Spirula spirula, that even approaches the belemnite morphology. Furthermore, whereas nautiloids and shelled coleoids were once widespread, from the Pliocene onwards they have become very restricted in distribution and ecology. The vast majority of living cephalopods are soft-bodied and shell-less: the squids and octopuses. Why should this be? What happened in the Miocene to make having a tradition cephalopod shell unviable after having been a pre-requisite to cephalopod success for 500 million years? One possible explanation is the evolution and radiation of echolocating whales and dolphins. Hollow, gas-filled shells produce very loud echoes, whereas soft tissues do not. Did whales and dolphins bring about the demise of the shelled cephalopods? Can we recognize species in the fossil record? A Caribbean gastropod case study Jonathan Todd and Timothy Rawlings Depít of Palaeontology, Natural History Museum, London, SW7 5BD. Within the context of adaptive radiations - focused upon here -there is a multitude of reasons why recognition of "true species" (biospecies equivalents) in the fossil record is necessary. If we are to assess intra-clade spatiotemporal diversity patterns, examine morphological changes associated with speciation and stasis, and evaluate the mechanisms underlying them, we need objectively delimited Recent biospecies in clades with an excellent fossil record. We can then use exactly the same morphological criteria to identify fossil and recent species. This is more challenging today than it was 20 years ago: molecular advances have revealed cryptic species to be ubiquitous in marine invertebrates. Can these genetic clades be delimited in fossil molluscs? Any study attempting to do this requires criteria to discriminate intraspecific from interspecific morphological variation. The Miocene to Recent Neotropical gastropod Polystira meets these criteria. We will explore morphological and molecular techniques by which we expect Polystira biospecies to be delimitable in the fossil record in our continuing study of this spectacular radiation. |
|