The Society's evening meeting, held jointly with the Linnean Society at their Burlington House rooms on 8th June 2000, was well received, with some keen questioning following the four talks. The President, Professor Robert Cameron, chaired the meeting. A wine reception followed the talks.
The Stylommatophora - before molecules Peter B Mordan. Mollusca Research Group, Department of Zoology, The Natural History Museum, Cromwell Road, London SW7 5BD, UK Email: pbm...@nhm.ac.uk Chris Wade and I, together with Bryan Clarke from Nottingham University, are working towards a molecular phylogeny of the stylommatophoran pulmonates. This presentation set the scene for Chris's talk by tracing the history of ideas on the deep-level relationships within the Stylommatophora up to the advent of molecular studies. The real breakthrough came with a short paper published by Henry Pilsbry exactly a century ago in 1900, entitled 'On the zoological position of Achatinella and Partula'. In this he defined three primary divisions of Stylommatophora based on differences in their pallial anatomy: the Orthurethra, the Sigmurethra and the Heterurethra. Later H. Burrington Baker added a fourth, the Mesurethra, comprising a small subset of Pilsbry's Sigmurethra. The 'Pilsbry-Baker' system, as it became known, remains the basis of all modern classifications. A comparison of the most recent classifications shows important areas of disagreement between them. All, however, agree as to the paraphyletic nature of Baker's Mesurethra. They also all accept the validity of the Orthurethra, as well as the primitive nature of the orthurethrous kidney, but several exclude the family Partulidae from this group.
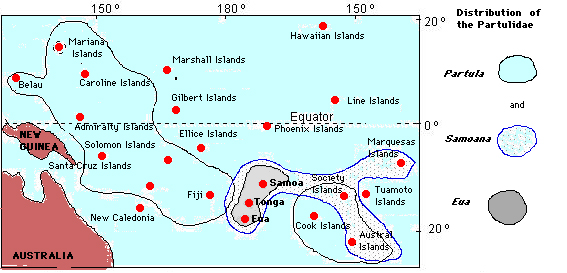
The Stylommatophora - a molecular phylogeny Christopher M. Wade1, Peter B. Mordan1 and Bryan C. Clarke2. 1Department of Zoology, The Natural History Museum, Cromwell Road, London SW7 5BD. 2Institute of Genetics, University of Nottingham, Queens Medical Centre, Clifton Boulevard, Nottingham NG7 2UH. We have undertaken the first large-scale molecular phylogenetic analysis of the Stylommatophora. Sequences of the ribosomal RNA gene-cluster were examined in 104 species of snails and slugs from 50 families, encompassing all the currently-recognised major groups. It allows an independent test of the present classification based on morphology. At the level of families our molecular phylogeny closely supports the current taxonomy, but the deep branches within the tree do not. Surprisingly, a single assemblage comprising the families Achatinidae, Subulinidae and Streptaxidae lies near the base of the tree, forming a sister group to all remaining stylommatophorans. This primary division into "achatinoid" and "non-achatinoid" groups is unexpected, and demands a radical reinterpretation of early stylommatophoran evolution. In particular, the Orthurethra appear to be relatively advanced within the "non-achatinoid clade", and broadly equivalent to other super-familial clusters. This suggests that supposedly primitive features such as the orthurethran kidney are derived. The molecular tree also indicates that the origin of the Stylommatophora is much earlier than the main period of their diversification. Convergent evolution in the family Partulidae Sara Goodacre, University of Nottingham. Current address: Cellular Genetics Group, Institute of Molecular Medicine, Headington, Oxford OX3 9DS (Sara...@molecular-medicine.oxford.ac.uk Partulid land snails of the tropical Pacific have evolved into an extraordinarily diverse array of different morphological, behavioural and ecological types that give excellent opportunities to study adaptation and speciation (Crampton 1932, Clarke and Murray 1969, Johnson et al 1993). There are three genera within the Partulidae: Partula, which is widespread on high islands of the Pacific from Belau in the west to the Austral Islands in the east, Samoana, which is found only on islands between Samoa in the central Pacific and the Marquesas in the west, and Eua, which is restricted to the Tongan and Samoan archipelagos.
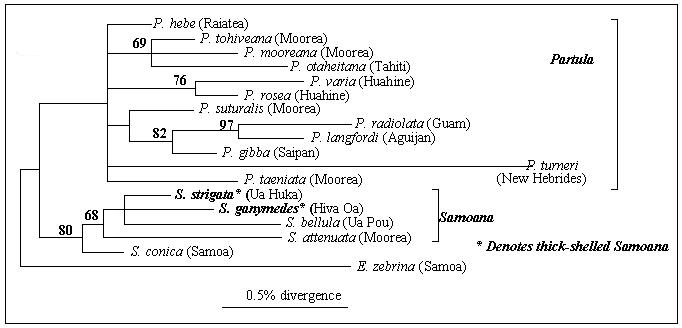
Species within each genus are broadly defined on the basis of reproductive anatomy, but there is also a suite of characteristics, including shell thickness, mantle pigmentation, tentacle length and type of mucus, that is loosely correlated with generic classification. Samoana from The Society Islands have thin, transparent shells, a pigmented mantle, long tentacles and sticky mucus, whereas Partula have thicker, pigmented shells, shorter tentacles and less sticky mucus. There are however some instances outside The Society Islands where Samoana resemble Partula in appearance. These Samoana, found in the Marquesas Islands, have thick, banded and pigmented shells, non-sticky mucus and short tentacles, although their reproductive anatomy appears to be characteristically Samoana-like. The apparently conflicting results for the Marquesan Samoana pose the question: Which characteristics are true indicators of the genus to which a species belongs? Either the current classification is wrong and the Samoana in question should be re-classified as Partula, or the suite of characteristics including shell, tentacle and mucus type has more than one origin. The present study uses DNA sequence data from a nuclear gene that encodes part of the ribosomal RNA (rRNA) protein translating machinery, to reconstruct evolutionary relationships between different partulid snails. rRNA genes have previously been used to investigate relationships at a deep level, such as between animal orders, but have also shown themselves to contain useful information about more recently diverged taxa (Hillis and Dixon 1991, Wade et al in manuscript). Sequences were obtained from 12 species of Partula, from Vanuatu, Guam and the Mariana Islands in the Pacific, and from Raiatea, Huahine, Moorea and Tahiti from the eastern Pacific. Sequences were also obtained from 5 species of Samoana, from Samoa in the central Pacific, Moorea in The Society Islands and the Marquesas Islands in the eastern Pacific. Eua zebrina from Samoa was also included in the analysis. The resulting evolutionary tree clearly separates the three genera from one another (see figure 2). Partula form a group separate to Samoana and Eua. Samoana also form a distinct group that includes both thin and thick-shelled representatives. The results agree well with the classification based on genital anatomy and with recent allozyme data, and show that the suite of characteristics incorporating shell thickness, mantle pigmentation, type of mucus and tentacle length, must have at least dual origins. The most likely explanation for morphological resemblances between Partula and Samoana is that they are the consequence of adaptive convergence. Repeated evolution of different morphological traits is perhaps not uncommon in land snails. Other characteristics that appear to have evolved more than once include the G-type clausial apparatus in the Clausiliidae, which protects against desiccation (Gittenberger and Shilthuizen 1996), the reduction of shell size in slug families (Tillier1989) and traits involved in the evolution of predation (Watson 1915).
In addition to resolving questions of taxonomy, reconstructing evolutionary relationships between species that are endemic to single islands may be informative about the order of colonisation. The route by which the Partulidae colonised the Pacific is obscure. They are not represented outside the pacific islands, and their closest continental relatives are unknown. It is most likely that they have their closest relatives in SE Asia or continental Australia (Gondwanaland) and colonised the pacific by trans-oceanic dispersal from west to east. The evolutionary tree presented in this study is consistent with colonisation by Samoana from west to east, although it provides little evidence for the direction of movement of Partula. Future studies using additional genes may help us to understand more about the manner in which partulids colonised the pacific. Further work may also refine our understanding of how such a diverse range of species originated, within which some suites of characteristics appear to have evolved more than once. References: 1. Goodacre, S. L., Wade, C. M. Molecular evolutionary relationships between partulid land snails of the Pacific - submitted to the Proceedings of the Royal Society, series B 2. Clarke, B., Murray, J. Ecological genetics and speciation in land snails of the genus Partula. Biol. J. Linn. Soc. 1, 21-42 3. Crampton, H. E. Studies on the variation, distribution, and evolution of the genus Partula. The species inhabiting Moorea. Carnegie Inst. Wash. Pub 410, 1-335 4. Gittenberger, E., Schiltzhuizen, M. 1996 Parallelism in the origin of the G-type clausial apparatus (Gastropoda, Pulmonata, Clausiliidae). In: Origin and Evolutionary radiation of the Mollusca ed. Taylor, J. D. Oxford University Press 5. Hillis, D. M., Dixon, M. T. 1991 Ribosomal DNA: molecular evolution and phylogenetic inference. Q. Rev. Biol.66, 411-453 6. Johnson, M. S., Murray, J., Clarke, B. The ecological genetics and adaptive radiation of Partula on Moorea. Oxford surveys in evolutionary biology 9, 167-238 7. Tillier, S. 1989 Comparative morphology, phylogeny and classification of land slugs and snails (Gastropoda: Pulmonata:Stylommatophora.) Malacologia 30, 1-303 8. Watson, H. 1915 Studies on the carnivorous slugs of South Africa, including a monograph of the genus Apera, and a discussion on the phylogeny of the Aperidae, Testacellidae and other agnathous Pulmonata. Annals of the Natal Museum 3, 107-267 A molecular eye on arionid slug relationshipsKarin Breugelmans & Thierry Backeljau. Royal Belgian Institute of Natural Sciences, Vautierstraat 29, B-1000 Brussels, Belgium. Email: TBac...@kbinirsnb.be Terrestrial slugs of the family Arionidae are widely distributed and several species are well-known agricultural pests. Particularly members of the genus Arion are notorious in this respect. It is therefore surprising that the taxonomy and phylogenetic relationships of these animals are still poorly understood. This is mainly due to the fact that Arion species are externally very similar and mainly differ by their colour. However, colour in terrestrial slugs is usually highly polymorphic and, worse, it often reflects environmental or physiological conditions (e.g. diet, humidity, age, ...) rather than taxonomic differences. Arionid anatomical features, on the other hand, often do provide species specific markers, but are at the same time usually very subtle and difficult to interpret, particularly with respect to their phylogenetic significance. As a consequence of all this, several 'familiar' Arion species appear to be in fact complexes of cryptic taxa (e.g. the Arion hortensis aggregate), while some recently described species do not merit a taxonomic separation at all (e.g. A. urbiae and A. anguloi). If one finally also realizes that arionid slugs may have variable and/or mixed breeding systems (outcrossing and selfing), then it may become clear why the taxonomy of these slugs is currently so confusing (Backeljau et al. 1996). In an attempt to clarify a number of these taxonomic and phylogenetic problems, we determined the complete nucleotide sequence of the nuclear ribosomal internal transcribed spacer I segment (ITS-1; ± 750 base pairs long) of more than 50 arionid taxa. The resulting ITS-1 alignments were then subjected to various tree reconstruction methods, including distance matrix methods, maximum parsimony and maximum likelihood. Tree stability was tested via bootstrap resampling. As 'reference' taxa (i.e. outgroups) we used three North American genera, viz. Ariolimax, Prophysaon and Hemphillia. The ITS-1 data analyses corroborated several current hypotheses and provided many new insights concerning the taxonomy and relationships of Arion species. For example the ITS-1 sequences supported the idea that A. intermedius belongs to the subgenus Kobeltia and hence should not be assigned to a separate subgenus (Microarion). Similarly, the data confirmed that A. urbiae and A. anguloi should not be considered as distinct species. On the other hand, the analyses also provided an illustration of how different alignment procedures (= homology determination) may produce very different, but significant tree topologies, leading to opposite conclusions! An important aspect of this kind of DNA work was also the possibility to include preserved museum specimens in the analyses. For example, we managed to extract DNA from an Azorean 'A. lusitanicus'-like slug collected in 1970 and preserved in 65% ethanol that was purchased in a local drugstore. For two A. distinctus specimens we extracted DNA from frozen albumen gland homogenates that were used for electrophoretic analyses, now more than 15 years ago (see Backeljau 1985, 1989). The first results of the DNA sequence analysis here presented will be published in the near future and currently we are extending our work to other genera such as Geomalacus and Ariolimax. References: 1. Backeljau, T. 1985. A preliminary account of species specific protein patterns in albumen gland extracts of Arion hortensis s.l. (Pulmonata, Arionidae). Basteria, 49: 11-17. 2. Backeljau, T. 1989. Electrophoresis of albumen gland proteins as a tool to elucidate taxonomic problems in the genus Arion (Gastropoda, Pulmonata). - Journal of Medical and Applied Malacology, 1: 29-41. 3. Backeljau, T., Winnepenninckx, B., Jordaens, K., De Wolf, H., Breugelmans, K., Parejo, C. & Rodriguez, T. 1996. Protein electrophoresis in arionid taxonomy. British Crop Protection Council Symposium Proceedings, 66: 21-28.
|
|